What are Tintinnids?
Species, Classification and Characteristics
Overview
Found in marine environments, tintinnids are protozoa belonging to the order Tintinnida. As compared to oligotrich ciliates and heterotrophic dinoflagellates among others that make up the microzooplankton population in marine systems, they are a minor group. However, they are also ubiquitous in this environment with a feeding behavior that sets them apart from the others.
According to studies, tintinnids are responsible for the consumption of the majority of the primary production in oceans across the globe and also serve as prey for a number of organisms (e.g. copepods).
Because of their unique morphological characteristics, researchers have been able to identify and distinguish well over 1000 species.
Some of the most common Tintinnid species include:
- Tintinnopsis beroidea
- Tintinnopsis radix
- Tintinnopsis fimbriata
- Tintinnopsis rapa
- Tintinnopsis parva
- Xystonella treforti
Classification
· Kingdom: Chromista - Consists of single-celled and multicellular eukaryotes possessing photosynthetic plastids
· Infrakingdom: Alveolata - Consists of protists belonging to the domain Eukarya
· Phylum: Ciliophora - Protozoa characterized by the presence hair-like organelles known as cilia (used for movement)
· Class: Spirotrichea - Ciliates characterized by the presence of an oral cavity. Being members of the phylum Ciliophora, these organisms also tend to have oral cilia
· Order: Tintinnida
 Tintinnids by Tintinnidguy / CC BY-SA (https://creativecommons.org/licenses/by-sa/4.0)")
Characteristics
Sexual and Asexual Reproduction
Binary fission
Like many other organisms, the growth rate of tintinnids has been associated with environmental conditions. During favorable conditions (characterized by increasing temperature and food abundance), the growth rate of tintinnids also increases. In the process, they are able to consume significant produce of marine phytoplankton.
With regards to reproduction, binary fission, a type of asexual reproduction, has been shown to be the typical mode of replication among Tintinnids. Following the division of cell components during the early stages of this process, lateral development of a new peristome is clearly evident. This is a clear indication that cell division has begun.
Here, the peristome belongs to one of the new daughter cells (known as the opisthe). This is then followed by the buildup of lorica building material (opaque granules) at the posterior part of the organism. This material then starts moving to the posterior part where they ultimately concentrate above the newly formed peristome.
Before the two cells separate (proter and opisthe), the adoral membrane of the opisthe (posterior cell of the two daughter cells) starts beating more actively than before, which helps in ejecting the other daughter cell (proter) as the two cells separate.
A few minutes after this separation, the proter starts to develop its own lorica (covering shell) while the other daughter cell retains the shell of the original parent cell.
This process may be represented as follows:
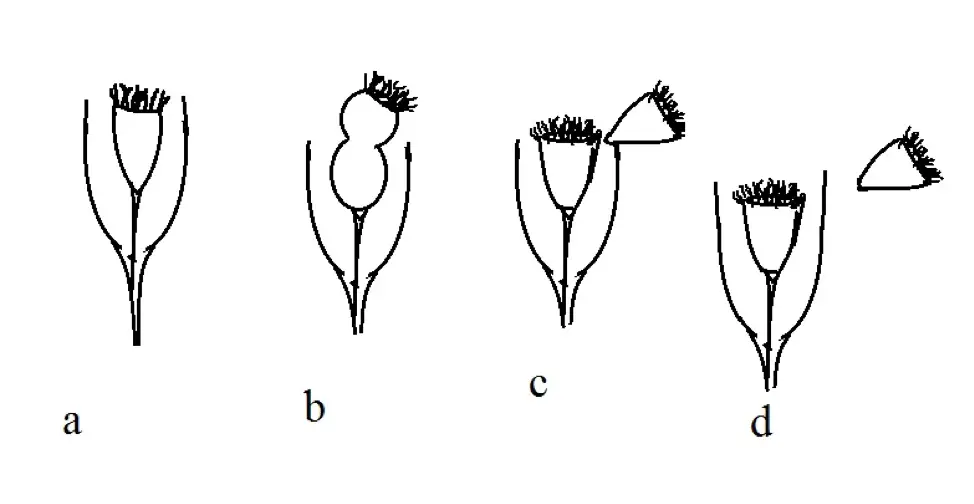
In this diagrammatic representation, the cell that separates and swims away is referred to as proter while the other daughter cell is referred to as opisthe.
Conjugation
While binary fission is the primary mode of reproduction, tintinnids have also been shown to reproduce through conjugation (a form of sexual reproduction).
Although it's common in tintinnid cultures, it's rare in nature. During conjugation, a microgamete is first produced post-laterally by the mother cell.
Once the gamete is produced (microgamete) a fracture occurs in the shell (lorica) allowing the gamete to be exposed to any receptive macrogamete (from a male tintinnid). This allows for gamete exchange between compatible cells.
* For the most part, sexual reproduction is induced by unfavorable environmental conditions (e.g. lack of food source).
Morphological Characteristics of Tintinnids
Within marine environments, Tintinnid species are diverse, ranging between 20 and 200um in size. Because they are relatively large in size, it's possible to view some of them using a magnifying glass. Compared to many other ciliates, one of their most defining characteristics is their shells (loricae).
While these structures can be found in some of the other planktons (e.g. pteropods), loricae found among tintinnid species are diverse with different forms between species.
When observed under the microscope, their shapes range from vase to bowl-like among others. They are closed/tapered at one end (posterior), but open at the oral end. This is not only important for feeding purposes, but also for cell division purposes.
* While a number of plankton possess shells, tintinnids are the only ciliates with this structure.
Because of the diversity in these shells, researchers have been able to differentiate among species. Shell characteristics have also been used to divide them into two main groups: Tintinnids with agglutinated shells and those with hyaline shells.
Whereas tintinnids with agglutinated shells (consisting of mineral particles) are common in coastal water, those with hyaline shells are generally found in open waters.
 while the brown life forms are tintinnids (grazers) by NOAA Photo Library:fish2778, https://creativecommons.org/licenses/by/2.0/ no changes made.")
%20Tintinnids%20by%20Tintinnidguy%20%2F%20CC%20BY-SA%20(https%3A%2F%2Fcreativecommons.org%2Flicenses%2Fby-sa%2F4.0)){kind=link}
%20while%20the%20brown%20life%20forms%20are%20tintinnids%20(grazers)%20by%20NOAA%20Photo%20Library%3Afish2778%2C%20https%3A%2F%2Fcreativecommons.org%2Flicenses%2Fby%2F2.0%2F%20no%20changes%20made.){kind=link}
Some of the main functions of lorica have been suggested to include:
· To provide protection from predators - where they also allow the organism to sink faster to evade predators
· Contribute to floatation
· Increase fluid motion during swimming - particularly among organisms with tubular shells
* Apart from the diversity in shell shapes, the contractile behavior of these shells is also one of the other defining characteristics.
The ciliate cell is itself attached to the lower part of the shell by a stalk-like structure referred to as the peduncle (contractile part of the cell). Whenever disturbed, the cell retracts and remains inside the shell. However, during feeding, it extends outwards through the open end where it's exposed to the surrounding environment.
At the anterior part of the cell, tintinnids have an oral opening/cavity surrounded by oral cilia. Apart from locomotion, these cilia are also used for feeding.
* During feeding, studies have shown the organism to use oral cilia to propel the cell to the anterior part of the shell (loricae) during feeding.
* In addition to oral cilia, the oral cavity is also surrounded by elongated structures known as tentaculoids (used to capture prey).
* Tintinnid cells occupy between 12 to 50 percent of the shell volume.
Habitat
As microzooplanktons, tintinnids are common in oceans across the world. Here, different species can be found in different environments with varying characteristics.
Whereas members of the genera Parafavella are often found in Arctic and Subarctic waters, members of the genera Laackmanniella are restricted to Subantarctic and Antarctic waters. In these environments, they feed on nanoplankton and picoplankton.
Despite being a minor group, their feeding behavior has been shown to dominate that of microzooplankton. Apart from phytoplankton, they can also feed on various organic matter and heterotrophic nanoflagellate.
They are also prey to a number of metazoan zooplankton like copepods. During the feeding process, oral cilia move in a manner that creates a current to suck in prey (in addition to aiding movement of the cell).
Return from Tintinnids to MicroscopeMaster home
References
J.A. Laybourn-Parry. (1992). Protozoan Plankton Ecology.
John R Dolan. (2012). Tintinntinid Ciliates: an Introduction and Overview. in Biology and Ecology of Tintinnid Ciliates: Models for Marine Plankton.
John R Dolan and Richard W. Pierce. (2012). The Biology and Ecology of Tintinnid Ciliates: Models for Marine Plankton.
Zhang, C. et al. (2017). Tintinnid community structure in the eastern equatorial Indian Ocean during the spring inter-monsoon period.
Links
http://aquaparadox.obs-vlfr.fr/html/PFD/2010/Dolan2010AP.pdf
https://sites.evergreen.edu/vms-spring/tintinnid-ciliate/
Find out how to advertise on MicroscopeMaster!


MicroscopeMaster.com is a participant in the Amazon Services LLC Associates Program, an affiliate advertising program designed to provide a means to earn fees by linking to Amazon.com and affiliated sites.
Recent Articles
-
Deltaproteobacteria - Examples and Characteristics
Nov 01, 22 04:44 PM
 Deltaproteobacteria is a large group (Class) of Gram-negative bacteria within the Phylum Proteobacteria. It consists of ecologically and metabolically diverse members. Read more here.
Deltaproteobacteria is a large group (Class) of Gram-negative bacteria within the Phylum Proteobacteria. It consists of ecologically and metabolically diverse members. Read more here. -
Chemoorganotrophs - Definition, and Examples
Oct 26, 22 05:01 PM
 Chemoorganotrophs also known as organotrophs, include organisms that obtain their energy from organic chemicals like glucose. This process is known as chemoorganotrophy. Read more here.
Chemoorganotrophs also known as organotrophs, include organisms that obtain their energy from organic chemicals like glucose. This process is known as chemoorganotrophy. Read more here. -
Betaproteobacteria – Examples, Characteristics and Function
Oct 25, 22 03:44 PM
 Betaproteobacteria is a heterogeneous group in the phylum Proteobacteria whose members can be found in a range of habitats from wastewater and hot springs to the Antarctic. Read more here.
Betaproteobacteria is a heterogeneous group in the phylum Proteobacteria whose members can be found in a range of habitats from wastewater and hot springs to the Antarctic. Read more here.